
Pająki mają długą historię, ale szczegóły ich ewolucji nie są do końca poznane. W 2018 roku opisano pajęczaka znalezionego w bursztynie datowanym na około 100 milionów lat. Okaz został znaleziony w birmańskim bursztynie i może stanowić klucz do poznania jednego z etapów ewolucji pająków. Zanim przejdziemy do dokładniejszego omawiania jego cech oraz cech poszczególnych grup pajęczaków, warto zerknąć na poniższy schemat przedstawiający ich wzajemne pokrewieństwa.
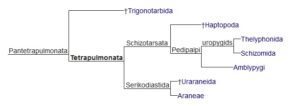
Fig. 1 | Drzewo filogenetyczne pajęczaków.
Dotychczas opisano ponad 47 tysięcy gatunków żyjących oraz ponad 1300 gatunków kopalnych pająków. Najstarsze szczątki tej grupy datowane są na późny karbon (około 305 mln lat temu). Wcześniejsze szczątki pająka z dewonu (380 mln lat temu) po późniejszej interpretacji okazały się wymarłym Uraraneida. Uraraneida są znane również z Permu (275 mln lat temu), przypominały pająki, lecz w przeciwieństwie do nich – posiadały długi witkowaty telson. Inną kluczową różnicą jest to, że Uraraneida mają brodawki przędne do produkcji nici jedwabiu, ale brakuje im kądziołków przędnych, które ułatwiają dokładne manipulowanie nicią.
Niedawno kilka paleozoicznych skamielin umieszczono blisko pajęczych przodków, gdyż posiadają one niektóre cechy charakterystyczne dla tej grupy. Uraraneida (dewon-perm) posiadały plezjomorficzny wydłużony telson i brodawki przędne, ale nie miały kądziołków przędnych. Szczegółowa analiza formalnie sytuowuje Uraraneida i Araneae w kladzie Serikodiastida. Inny pająko-kształtny kopalny takson – Idmonarachne brasieri posiadał zarówno kądziołki przędne jak i telson. Te cechy sugerują, że klad ten jest bliżej spokrewniony z Araneae niż z Uraraneida. Posiadał również podzielony odwłok na tergity jak u członków wymarłej grupy Trigonotarbida.
Analiza filogenetyczna często sugerowała, że pająki są siostrzaną grupą dla innych pajęczaków: tępoodwłokowców (Amblypygi), biczykoodwłokowców (Thelyphonida) i rozłupnogłowców (Schizomida). Podobieństwa pająków z pozostałymi Pantetrapulmonata to posiadanie dwóch par płucotchawek. Biczykoodwłokowce i rozłupnogłowce posiadają wydłużony telson, charakterystyczny również dla grup nieposiadających dwóch par płucotchawek – rząd głaszczkochody (Palpigradi), który zachował kilka plezjomorfii wśród pajęczaków (cech odziedziczonych po wspólnych przodkach). Kluczową autopomorfią pająków podkreślającą ich wysoki stopień rozwoju są gruczoły przędne zakończone brodawkami na kądziołkach przędnych, szczękoczułki z gruczołami jadowymi i narządy kopulacyjne samców na nogogłaszczkach. Pająki są tradycyjnie podzielone na dwa podrzędy. U Mesothelae zachowały się jeszcze segmenty na odwłoku, a kądziołki przędne są położone na spodniej stronie w połowie długości odwłoka. Opisthothelae (Mygalomorphae i Araneomorphae) nie mają w większości zewnętrznych dowodów na segmentację opisthosomalną, a kądziołki przędne przesunęły się w kierunku tylnej części ciała.

Fig. 2 | Holotyp Chimerarachne yingi. a,c, Cały okaz od grzbietowej (a) i brzusznej strony (c). b,d, Rysunki interpretacyjne odpowiadające zdjęciom a i c. IIV numeracja odnóży krocznych. ALS, (anterior lateral spinneret) przednie boczne kądziołki przędne; car, karapaks; ch, chelicery; cx, biodro; fe, udo; lb, warga; mt, przedstopie; op, wieczko; p, stylik; pa, rzepka; Pd, nogogłaszczek; PLS, (posterior lateral spinneret) tylne boczne kądziołki przędne; pyg, pygidium; st, sternum; t, trichobothrium; ta, tarsus; ti, tibia; tr, krętarz. Jednostka skali: 1!mm.
Tak jak Uraraneida, Chimerarachne posiada segmentowany odwłok zakończony biczykiem przypominającym telson. Prócz tego posiada również dwie synapomorfie występujące u współczesnych pająków: narządy kopulacyjne samców na nogogłaszczkach do przekazywania spermy oraz dobrze wykształcone kądziołki przędne (jak u współczesnych Mesothelae). Te cechy sprawiają, że gatunek Chimerarachne yingi ma wspólne cechy zarówno z Uraraneidea jak i Araneae. Oznacza to, że ten nowy materiał kopalny stanowi początek gałęzi ewolucji pająków.
Materiał badawczy
Holotyp i paratyp zostały zdeponowane w Instytucie Geologii i Paleontologii w Nanjing, Chińska Akademia Nauk. Oba okazy są samcami. Długość ciała bez telsona: 2,42 mm i 2,80 mm, długość witkowatego telsona: 2,86 mm (drugi nie zachował się w całości).
Etymologia nazwy
Rodzaj: Chimerarachne (chimera: ona-koza) – z greckiej mitologii, hybrydowe stworzenie złożone z części dwóch lub większej ilości zwierząt; i z języka greckiego (Ἀράχνη, Aráchnē) (arachne: pająk) – postać z greckiej mitologii tkająca nici. Gatunek: Chimerarachne yingi – nazwa honorująca patronat Yanling Ying.

Fig. 3 | Paratyp Cimerarachne yingi, a,c, Cały okaz od grzbietowej (a) i brzusznej strony (c). b,d, Rysunki interpretacyjne odpowiadające zdjęciom a i c. IV numeracja odnóży krocznych. ALS, (anterior lateral spinneret) przednie boczne kądziołki przędne; car, karapaks; ch, chelicery; cx, biodro; fe, udo; lb, warga; mt, przedstopie; op, wieczko; p, stylik; pa, rzepka; Pd, nogogłaszczek; PLS, (posterior lateral spinneret) tylne boczne kądziołki przędne; pyg, pygidium; st, sternum; t, trichobothrium; ta, tarsus; ti, tibia; tr, krętarz. Jednostka skali: 1!mm.
Dyskusja
Chimerarachne yingi jest kluczem do zrozumienia powiązań pająków z innymi grupami pajęczaków. Jego charakterystyczną cechą jest witkowaty telson, ciało podzielone na głowotułów i odwłok (połączone wąskim stylikiem). Głowotułów nie jest podzielony. Znajdują się na nim oczy ułożone w dwóch rzędach, zarówno boczne jak i środkowe znajdują się na wzgórkach. Chelicery są takie jak u pająków, w sztyletowatym kształcie. Są ustawione w pozycji prostopadłej do osi ciała, pod kątem 45°C. Krawędzie zębów są nagie. Nogogłaszczki mają kształt nóg krocznych. U samców są zakończone rozwidloną stopą, z cymbium składającym się z bulba i embolusa. Opistosoma jest podzielona na segmenty i kończy się czterosegmentowym zaodwłokiem, na którym osadzony jest wić, dłuższa od ciała.
Jeśli chodzi o stronę brzuszną, trzeci segment stanowi podstawę dla pary długich wielosegmentowych kądziołków przędnych (odpowiadających przednim kądziołkom bocznym u pająków) oraz mniej rozwiniętych wzgórków z brodawkami przędnymi (które prawdopodobnie odpowiadają środkowej parze kądziołkąw przędnych). Na czwartym segmencie opistosomy znajduje się kolejna para wielosegmentowych kądziołków przędnych (odpowiadające tylnym kądziołkom bocznym u pająków).
Zatem posiadanie witkowatego telsonu jest przypuszczalnie plezjomorficzną cechą. Niezwykłym aspektem tego odkrycia jest to, że zwierzęta z takim planem budowy ciała były nadal obecne w późnym mezozoiku. Chimerarachne yingi żył obok kredowych pająków należących do wielu współczesnych rodzin. Następstwem tego jest to, że nowe skamieliny nie mogą stanowić wczesnego ogniwa ewolucji dla współczesnych pająków, które (jak wspomniano we wstępnie) były reprezentowane już w karbonie. Chimerarachne yingi prawdopodobnie reprezentuje grupę, która wyodrębniła się z wcześniejszej (być może paleozoicznego) radiacji Pantetrapulmonata.

Fig. 4 | Morfologiczne szczegóły i rekonstrukcja. a,b, Rekonstrukcja grzbietowej (a) i brzusznej strony (b). c, oczy. d, szczękoczułki. e, f, Brzuszna strona narządów kopulacyjnych. g, Kądziołki przędne. h, Obraz spodu w konfokalnym mikroskopie skaningowym . i,j, Pazury na II odnóżu krocznym. k, Witkowaty telson. ap, apofiza; ALS, przednie boczne kądziołki przędne; ch, chelicery; em, embolus; ey, oczy; mc, środkowy pazur; op, wieczko; pc, boczne pazury; PLS, tylne boczne kądziołki przędne; pu, przylga; sp, brodawki przędne; ta, stopa. Jednostka skali: 1!mm w a i b; 0.2!mm w eh i k; 0.1!mm w c i d; 0.05!mm w i oraz j.
Bursztynowe inkluzje są również bardzo interesujące dlatego, że ukazują kluczowe informacje na temat zmian w obrębie gałęzi ewolucyjnej pająków i prawdopodobną morfologię przodków Araneae. Oczy w skamielinie wydają się nie znajdować na jednym guzku, jak na przykład u Mesothele. Zamiast tego składają się z pary środkowych oczu i dwóch bocznych guzków z różną liczbą soczewek. Taka budowa jest charakterystyczna dla biczykoodwłokowców, tępoodwłokowców i niektórych pająków należących do Mygalomorphae (nadrodzina Atypoidea: rodzina Hypochilidae) – które oddzieliły się od reszty we wczesnym etapie ewolucji pająków.
Chelicery Chimerarachne yingi są zbudowane jak u pająków, z nagim (bezwłosym) zębem. Stan zachowania szczątków nie pozwala sprawdzić czy ząb ma otwór dla gruczołu jadowego. Kluczową cechą w nowych skamielinach jest zmiana nogogłaszczek w to, co możemy zinterpretować jako narząd kopulacyjny, przy czym na stopie znajduje się cymbium ze zgrubieniem (prawdopodobnie zbiorniczek do przeniesienia spermy). Głęboko rozwidlona natura stopy / cymbium w nowej skamielinie, jest unikalna i jest potencjalną cechą diagnostyczną dla rodzaju. Należy jednak zauważyć, że u niektórych pająków z grupy Mygalomorphae stopa jest krótka i składa się z dwóch podobnych zaokrąglonych płatów. Zauważamy, że nogogłaszczki Chimerarachne yingi są bardziej podobne do tych u Mygalomorphae niż u Mesothelae. To może nasuwać pytanie, czy budowa narządu kopulacyjnego u Mygalomorphae jest bliższa cesze plezjomorficznej niż bardziej złożone strukturalnie narządy Mesothelae? Na przykład, jedno z przeprowadzonych badań pokazało, że proste narządy kopulacyjne Mygalomorphae zawierały wtórnie scalone elementy. Obecny materiał nie jest zachowany wystarczająco dobrze, aby odpowiedzieć na to pytanie, ale przyszłe znaleziska mogą się okazać bardzo pommocne.

Fig. 5 | a, Cymbium w confokalnym mikroskopie skaningowym. b, Odnóże II stopa i pazur. Widoczne ostatnie pięć trichobothrii na końcowej części stopy II. c, Prawe odnóże kroczne IV z pazurami. d, Lewe odnóże kroczne IV z pazurami. e, Stylik od strony brzusznej. f, Odwłok od strony brzusznej. g, Odwłok od strony grzbietowej. h, Witkowaty telson. i, Głowotułów od strony grzbietowej. j, Nogogłaszczki od strony brzusznej. Jednostka skali: 0.2!mm w f, g, i oraz j; 0.1!mm w a, d, e i h; 0.05!mm w b i c.
Być może najbardziej interesującym aspektem nowych skamielin jest pozycja i ułożenie kądziołków przędnych. To może sugerować, że pierwotną funkcją pajęczego jedwabiu nie było budowanie sieci, lecz pakowanie jaj. Mesothelae, jako siostrzana grupa wszystkich innych pająków zachowująca plezjomorficzne cechy (takie jak segmentacja) mogą wesprzeć tą teorię. Swoje kądziołki przędne mają umieszczone na środku brzusznej strony odwłoka w pobliżu otworu narządów płciowych. Kądziołki przędne przesunięte ku tyłowi to tradycyjna synapomorfia Opisthothelae. Chimerarachne yingi, jako potencjalna grupa wyjściowa dla wszystkich żywych pająków, podważa ten scenariusz, posiadając kądziołki przędne przesunięte ku tyłowi. Innym powszechnym założeniem jest to, że pająki pierwotnie miały cztery pary kądziołków przędnych: przednie środkowe, przednie boczne oraz tylne środkowe i tylne boczne. U Chimerarachne yingi zarówno przednie boczne, jak i tylne boczne kądziołki są dobrze rozwinięte i składają się z wielu krótkich części (pseudosegmentów). Są one podobne do kądziołków przędnych współczesnych Mesothelae, ale różnią się posiadaniem dwóch lub trzech dużych części u podstawy, wychodzących ze sklerotyzowanych płatów brzusznych. U Mesothele jest tylko jeden lub dwa duże części podstawowe i brak płatów brzusznych związanych z kądziołkami przędnymi. Tam, gdzie spodziewalibyśmy się w nowej skamielinie przednich środkowych kądziołków przędnych, jest tylko jedna para wzgórków z brodawkami przędnymi, podczas gdy tylne środkowe kądziołki wydają się być całkowicie nieobecne.
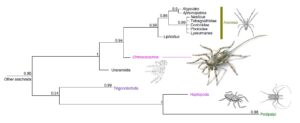
Fig. 6 | Relacje filogenetyczne. Kladogram zgodności (analiza Bayesowska przy silnym wsparciu) pokazuje pozycję Chimerarachne yingi wewnątrz gałęzi Pantetrapulmonata u pajęczaków, która obejmuje pająki I ich najbliższych krewnych. Zarówno Attercopus jak i Permarachne są niekompletne, a analizowanie ich indywidualnie powoduje załamanie się drzewa, dlatego do analizy połączyliśmy dwa rodzaje w jedną grupę Uraraneida.
Porównanie grup zewnętrznych pokazuje, że zachowanie kądziołków przędnych powinno być plezjomorficzne, ale krewne pająków, takie jak Uraraneida nie mają przydatków opisthosomalnych. Autorzy postulowali że u pająków może istnieć mechanizm genetyczny reaktywujący rozwój (utraconych) przydatków, umożliwiający ewolucję ruchomych kądziołków przędnych, które ułatwiają bardziej precyzyjną manipulację jedwabnymi nićmi. Chimerarachne yingi zachowuje tylko część przewidywanego podstawowego wzoru dla pająków, pozostawiając pytanie, czy środkowe kądziołki przędne w bursztynowych skamielinach są w trakcie formowania lub redukcji. Utrata skleritów brzusznych związanych z kądziołkami przędnymi na segmentach opistosforalnych 3 i 4 wyraźnie różnicuje wszystkie współczesne pająki od nowych skamielin. Obecność tych skleritów u Chimerarachne może poprzeć hipotezę, że to odzwierciedla plezjomorficzną anatomię brzuszną.
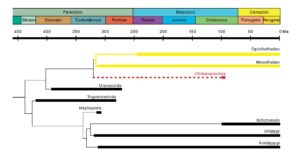
Fig. 7 | Historia ewolucji pająków i ich relacje z innymi pajęczakami. Grube linie wskazują znany zakres zapisu kopalnego. Gałąź reprezentująca współczesne pająki jest żółta. Gałąź Chimerarachne jest czerwona.
Czy Chimerarachne yingi jest pająkiem? Konsekwentna rekonstrukcja filogenetyczna ukazuje go w grupie z Uraraneida, Araneae i Idmonarachne, ale różne techniki analizy dają różne wewnętrzne topologie. Pod parsymonią (najbardziej prosty i oszczędny przebieg zmian) o równej wadze, ścisłe drzewo sugeruje, że nowy gatunek stanowi grupę siostrzaną dla Uraraneida. Analiza oparta na implikowanej wadze (cechy mające wspólne pochodzenie posiadają największą wagę) umieszcza go jako siostrzaną grupę wszystkich zachowanych pająków, a alternatywna analiza statystyczna opierająca się na twierdzeniu Bayesa ukazuje Uraraneida, Araneae, Idmonarachne i Chimerarachne w politomii. Ta niestabilność może wynikać z kilku kluczowych cech, takich jak ułożenie oczu i wszelkie modyfikacje męskich narządów kopulacyjnych, które są nieznane zarówno u Uraraneida, jak i u Idmonarache. Ten problem jest szczególnie dotkliwy u Idmonarache, gdzie zachowana morfologia jest zagadkowa. Aby przetestować wpływ tej skamieliny na wyniki, wykluczyliśmy Idmonarachne z analizy i uzyskaliśmy polimorfię u Uraraneida, pająków i Chimerarache przy równomiernej parsymonii oraz Chimerarachne z grupą pająków (w szczególności siostrzaną grupą do Mesothelae) pod analizą implikowanej wagi. Nasze preferowane drzewo opiera się na analizie bayesowskiej (bez Idmonarachne), która daje filogenezę grup (Uraraneida (Chimerarachne (Mesothelae + Opisthothelae))) przy silnym wsparciu. Sugerujemy, że załamanie się tego regionu drzewa, gdzie jest uwzględnione Idmonarachne prawdopodobnie odzwierciedla rzeczywistą kruchość tej metody. Kiedy uwzględnimy wszystkie końcowe taksony do analizy Bayesowskiej, to niektóre z nich mogą wymagać bardziej czułej analizy.
Wnioski
Niezależnie od dokładnych związków między grupami, dochodzimy do wniosku, że Chimerarachne ujawnia stopień organizacji, kiedy to pająki lub jeden z ich najbliższych krewnych rozwinęły męski narząd kopulacyjny na nogogłaszczkach i przynajmniej część nowoczesnego aparatu przędnego, ale zachowały pierwotny witkowaty twór podobny do telsonu. Zatem musiało istnieć kontinuum redukcji telsonu i rozwój innych cech w kierunku współczesnych pająków. I teraz nasuwa się pytanie – czy Araneae sensu stricto powinny być zdefiniowane przez utratę telsonu lub przez pojawienie się kądziołków przędnych i / lub męskich narządów kopulacyjnych na nogogłaszczkach?
AngelFresh
Źródło:
Wang B., Dunlop J. A., Selden P. A., Garwood R. J., Shear W. A., Müller P. and Lei X. 2018. Cretaceous arachnid Chimerarachne yingi gen. et sp. nov. illuminates spider origins. Nature Ecology & Evolution. DOI: 10.1038/s41559-017-0449-3
Liczba wyświetleń: 23










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}